Куда идем: к экокатастрофе или экореволюции?
скачать Автор: Зубаков В. А. - подписаться на статьи автора
Журнал: Философия и общество. Выпуск №6/1998 - подписаться на статьи журнала
Я смотрю на значение философии в развитии знания совсем иначе, чем большинство натуралистов, и придаю ей огромное плодотворное значение... Философия всегда заключает зародыши, иногда даже предвосхищает целые об ласти будущего развития науки...
В. И. Вернадский
ЧАСТЬ II . ОТКУДА МЫ? ФИЛОСОФИЯ ЭВОЛЮЦИИ11. Вопросы к размышлению
На вопрос «Где мы?» (человечество в целом) в первой части статьи был дан ответ – мы прошли половину пути от глобального экологического кризиса (ГЭК) к тотальной экокатастрофе (ТЭК), и ЕСЛИ стратегия развития цивилизации радикально не изменится, то ТЭК может разразиться в любое время, начиная с середины следующего столетия, возможно, и раньше. Неодолимое стремление потреблять все больше привело человечество к краю бездны. Самое страшное то, что 95% (по А. В. Яблокову) населения мира и его политической элиты НЕ понимает последствий ТЭК для себя и своих потомков.
Может ли человечество выжить в этой ситуации – центральный вопрос статьи. Может, ЕСЛИ успеет экологически прозреть. На это «прозрение» у нас есть 10, максимум 15–20 лет. Следовательно, оно может свершиться только в форме глобального ЭКОЛОГИЧЕСКОГО ВСЕОБУЧА. В процессе его человечество должно осмыслить главные итоги эволюции или, по крайней мере, понять ее логику и в соответствии с ней разработать новую СТРАТЕГИЮ ЖИЗНИ. Одним из главных предметов эковсеобуча должна стать философия эволюции, которая в отличие от науки об эволюции – синергетики – может быть доступна всем, от малого до старого.
Для того чтобы читателю было легче войти в круг философских проблем эволюции, сформулирую здесь несколько «вечных» вопросов к размышлению, субъективные ответы на которые, может быть , я сумею дать по ходу обсуждения. Сама постановка этих вопросов должна настроить читателя на критический диалог и дискуссию, в расчете на которую и писалась эта статья.
Итак, 1. Что такое эволюция? Я имею в виду не принятое в разговорной лексике противопоставление эволюции как плавного развития революции как скачку в развитии, а научное содержание термина. В него вкладывается сейчас два понятия – узкое, биологическое, и широкое, мировоззренческое. В первом эволюция понимается как синоним дарвинизма, как необратимое развитие живой природы, генетического состава популяций, возникновение и вымирание видов, преобразование биоценозов и биосферы в целом 2 . Во втором, говоря словами Р. С. Карпинской 3 , принцип биологического эволюционизма, сохраняя свою дарвиновскую сущность, «очеловечивается» и космологизируется. В рамках синергетики как науки о саморазвитии термин «эволюция» становится символом направленного усложняющегося саморазвития , в котором реализуются уже не три, а все четыре закона диалектики, включая Закон «развития через напряжение» (гистерезиса плотности массы) 4 . Такая эволюция в применении к Вселенной оказывается процессом более биологическим , чем физическим, представляя чередование относительно длинных плавных этапов развития с короткими взрывоподобными скачками–кризисами и революциями, ныне именуемыми, по И. Пригожину 5 , точками бифуркации (ТБ). Таким образом, кризисы и революции, а не просто переход количества в качество и борьба противоположностей, играют в эволюции роль кнута, которым процесс саморазвития направляется на более высокую в информационном плане ступень развития. Существенно, что эволюция имеет место только в неустойчивых системах и определяется вероятностными закономерностями.
2. Можно ли считать эволюцию направленной и что это означает? Иными словами, есть ли «логика эволюции»? По этому вопросу существует два мнения: а) эволюция определяется действием случайных факторов и потому непредсказуема в принципе, б) эволюция не случайна и не просто направленна, а целенаправленна, возможно, даже запрограммирована, а следовательно, траектории будущего могут исследоваться как вероятностные. Как будет показано ниже, анализ эволюции Земли и жизни на ней свидетельствует в пользу наличия закономерностей эволюции. А из этого следует, что свобода выбора стратегии по отношению к природе определяется мерой осознания нашего будущего.
3. Были ли ГЭК и ТЭК в прошлом, и чем они заканчивались? Это один из самых важных, если не главный из вопросов, потому что в зависимости от ответа на него определяется методика прогнозирования будущего. Мнения ученых-экологов по этому вопросу диаметрально расходятся. По мнению одних, опыт истории говорит о множестве глобальных кризисов и катастроф в прошлом. При этом палеоаналогом современного ГЭК является, по их мнению, в частности Н. Н. Моисеева 6 , Н. Ф. Реймерса 7 , С. И. Барановского и др. 8 , позднепалеолитический кризис. Следовательно, неолитическая революция может быть принята за палеоаналог выхода из современного ГЭК. Альтернативное мнение высказано В. Г. Горшковым 9 и группой ученых во главе с председателем Комитета РФ по экологии В. И. Даниловым-Данильяном 10 . Они утверждают, что никаких ГЭК, а тем более ТЭК в истории биосферы до сих пор не было, а следовательно, не может быть и никаких аналогов для поиска выхода из современного ГЭК. Я придерживаюсь третьей точки зрения, а именно, что первой задачей исторической геоэкологии является классификация экокризисов и экокатастроф прошлого на локальные, региональные, зональные и глобальные. Позднепалеолитический кризис, вызванный резким потеплением климата высоких широт и вымиранием в связи с этим мамонтового биогеоценоза, был чисто зональным событием. Современному же ГЭК в прошлом находится только один аналог – прерифейский кризис, бывший 2 млрд л. н. Причины и последствия его будут подробно рассмотрены ниже.
4. Что такое жизнь? Как уже указывалось в первой части статьи, на этот вопрос едва ли кто-нибудь сейчас может дать исчерпывающий ответ. В. И. Вернадский предпочитал даже говорить не о жизни, а о «живом веществе» 11 как совокупности организмов и о «биологическом пространстве-времени», отмеченном асимметрией и выражающемся в смене его единиц (поколений, видов, родов, типов и др.). Еще в 1931 г. он поставил «самый гениальный», по мнению Г. В. Гегамяна 12 , вопрос XX века – о возможной биологической структуре всего мироздания. Развивая эту мысль В. И. Вернадского, Г. В. Гегамян вводит понятие «поле живого вещества», включающее иерархию полей от поля биополимера и живой клетки до поля Вселенной. Сходная мысль развивается Дж. Тленном 13 , который принимает жизнь за пятую силу бытия. В. И. Гольданский и др. 14 также считают, что жизнь не могла возникнуть эволюционным путем, так как в этом случае существовали бы лево- и правостереоизомерные ее формы, и она являлась бы фундаментальным свойством Вселенной (а не простой комбинацией молекул, как традиционно считается). Теоретически сейчас предполагаются разные формы жизни – техножизнь, солитонно-голографи-ческая и др. Но практически мы знаем пока только одну форму – белково-нуклеиновую, функционирующую на Земле на базе обмена веществ, энергии и информации в процессах метаболизма.
5. Как появилась жизнь на Земле? Имеющиеся ответы, а их множество, можно разделить на две крайние позиции: а) в результате самозарождения в ходе длительной, не менее 2 млрд лет, химической эволюции. Так утверждает теория, развитая А. И. Опариным 15 , Г. Юри, Дж. Берналом 16 и др., до сих пор доминирующая в науке; б) в результате панспермии, т. е. переноса зародышей жизни из Космоса. Она была сформулирована Сванте Аррениусом и поддержана В. И. Вернадским. Сторонники ее (Фред Хойл и др.) доказывают, что 80% частиц межзвездной пыли будто бы представлены клетками бактерий и диатомовых водорослей. Согласно этой гипотезе, жизнь вечна и на Земле была, по Ф. Крику и Л. Оргелу, «засеяна» искусственно представителями внеземной цивилизации 17 . К кометному «засеву» жизни приходят также Г. В. Двойрин 18 и ряд других исследователей.
6. Является ли земная биота (и Земля) организмом? Этот вопрос был поставлен еще сподвижником Ч. Дарвина Г. Хаксли, в осторожной форме поддержан В. И. Вернадским и однозначно решен английским естествоиспытателем Дж. Лавлоком в его теории Геи 19 . Но М. М. Камшилов 20 и В. Г. Горшков 21 , признавая ранг организма за биоценозами, такую интерпретацию для Земли в целом отвергают. Автор, следуя В. А. Шухову и Г. Н. Хону 22 , считает, что организмом можно считать любую колебательную систему с собственным временем, которая «выбирает» варианты взаимодействия с окружающей средой. Ниже будет показано, что биосфера, «выбирая» 2 млрд лет назад переход от восстановительной стратегии взаимодействия со средой к окислительной, по сути самой своей эволюцией продемонстрировала свою принадлежность к разряду космического организма.
7. Что такое информация, является ли она атрибутом только живого, как многие считают, или всей Вселенной? По сути, понятие информации, противопоставленное Н. Винером понятию энтропии, является сейчас синонимом теории связи и управления. По Э. Шеннону и И. Р. Пригожину, информация – это то, что оставляет о себе активную «память» в веществе 23 . По определению президента Международной академии информациологии И. И. Юзвишина, «информация – это всеобщий единый законопроцесс фундаментальных отношений, связей, взаимодействий и взаимозависимостей энергии, движения, массы и антимассы микро- и макроструктур Вселенной. Мир информационен, Вселенная – информационна; первичное – информация, вторичное – материя. Не бытие определяет сознание, а информация (сознание) определяет бытие» 24 .
8. Как понимать время в эволюции? Как известно, существуют две трактовки этого фундаментального понятия: а) субстанциональная, как параметра измерения – «абсолютное», или физическое время дления (И. Ньютон, Н. А. Козырев) и б) реляционная, как отношения событий – «относительное» время (Г. Лейбниц, А. Эйнштейн). В первой время отделено от пространства, во второй вводится дополнительное понятие пространство-время. Вариантом второй трактовки является, в частности, понятие биологическое пространство-время, введенное В. И. Вернадским. Обзор современного состояния этого вопроса дан Л. С. Шихобаловым и А. А. Вакуленко и др. 25 Развивая идеи Н. А. Козырева 26 о ходе времени и плотности времени, они считают, что именно стрела времени является организующим фактором всей эволюции, т. е. несет негэнтропию. В построениях этих авторов остается много неясного. Нет, в частности, корреляции с идеями И. Пригожина и И. Стенгерса 27 о темпоральности как стреле времени и темпорализованном пространстве, оставленном асимметричными событиями.
2. Пришиты темпоральной периодизации эволюции Земли
Перечисленные выше «вопросы к размышлению», надеюсь, настроили читателя на критический диалог с автором в его поиске нового видения истории Земли и Общества в духе идей В. И. Вернадского и И. Р. Пригожина. Поэтому, отойдя от традиционной для науки хроностратиграфической шкалы описания истории Земли на эры и периоды, попробуем теперь разделить ее на «этапы взросления биосферы» (табл. 1). Подобно тому как жизнь каждого человека разграничивается на этапы такими рубежами, как «начал ходить», «начал говорить», вступил в половую зрелость, вступил в климакс, так и эволюция Земли как поля живого вещества может быть разделена на ряд возрастных этапов четкими рубежами–бифуркациями. Автором был разработан макет темпоральной (от лат. tempus – время) периодизации истории Земли и телеостра-тиграфической (от греч. teleos – цель) классификации отложений с использованием четырех таксонов (табл. 1). Описание истории Земли в предлагаемой публикации является элементом исторической геоэкологии – новой научной дисциплины, лежащей на стыке геологии, биологии, археологии, истории, экономики, социологии и философии эволюции.
Таблица 1. Таксоны темпоральной периодизации истории Земли как единицы бионоогенического пространства-времени
|
Темпоральные единицы времени (лат. tempts – время) |
Телеостратиграфические единицы отложений (греч. teleos – цель, them – объект обсуждения) |
|
Сферотемпа (греч. сфайра – шар) |
Сферотемп |
|
Гайятемп {греч. Гея – Земля) |
Гайятемп |
|
Генотемп (греч./лат. ген – родить) |
Генотемп |
|
Гемеротемп (греч. емер – день) |
Гемеротемп |
Под философией эволюции я понимаю такую часть философии, которая занимается осмыслением хода истории Земли и биосферы как дома человечества и поиском возможных закономерностей в развитии взаимоотношений человечества с природной средой.
Введение четырех таксонов в темпоральное описание истории Земли как организма означает не что иное как классификацию экологических кризисов и катастроф на локальные, региональные, зональные и глобальные кризисно-переходные рубежи, о которых шла речь выше (в разделе 1.3). Такая классификация позволила прийти к обшей схеме темпоральной периодизации эволюции Земли, изображенной в табл. 2. В правой ее колонке с помощью традиционных индексов указано соответствие объемов темпоральных единиц подразделениям всем известной геоисторической (хроностратиграфической) шкалы.
Ниже по необходимости кратко, даже тезисно, опишем главные этапы роста Геи как космического суперорганизма (табл. 2). Наш обзор представляет эмпирическое обобщение современных данных, нередко еще не получивших теоретического истолкования профессионалов. Целью его является общее философское осмысление совокупности данных.
Таблица 2. Темпоральная периодизация эволюции Земли
|
Сферотемп |
Гайятемп |
Генотемп |
Гемеротемп |
Лет назад |
эры |
|
III Ноосферотемп |
Техногайя или экогайя – выбор |
|
|
|
Голоцен |
|
Современный экологический кризис |
| ||||
|
Социогайя |
Графоген – цивилизация |
Инфогемер |
10 | ||
|
Механогемер |
200 | ||||
|
Агрогемер |
5–8 тыс | ||||
|
Эпоген – социализация |
Териогемер |
50–100 тыс |
Плейстоцен | ||
|
Каугемер |
1,75 млн | ||||
|
Афрогемер |
2,3–2,0 млн | ||||
|
|
Сексуальная (социальная) бифуркация |
2,3 млн |
Кайнозой | ||
|
II Биосферотемп |
Оксигайя |
Эстеген – цефализация |
Рамогемер |
40 млн | |
|
Хтоногемер |
450–400 млн |
Мезозой | |||
|
Пноеген – цитоколонизация |
Склерогемер |
570 млн |
Палеозой | ||
|
Малакогемер |
1 млрд |
Рифей | |||
|
Протистогемер |
1,7–1,6 млрд | ||||
|
|
Перифейский экологический кризис |
2–1,7 млрд |
| ||
|
|
Антракоогайя |
Фотоген |
Сидерогемер |
2,3 млрд |
Протерозой |
|
Цианогемер |
2,8 млрд | ||||
|
Нефоген |
|
3,8 млрд |
Архей | ||
|
|
Генетическая (хиральная) бифуркация |
4,0 млрд |
| ||
|
I Сейсмосферотемп |
Пирогайя |
|
|
4,0–4,6 млрд |
Катархей |
|
Кризис Роша | |||||
|
Гаплогайя | |||||
|
|
Планетообразование |
|
|
4,67 млрд |
|
3. Этап добиологической эволюции Земли – сейсмосферотемп
Разделение истории Земли именно на три сферотемпа (табл. 2) отражает бесспорный эмпирический вывод о направленности ее эволюции. Действительно, примерно до 4 млрд лет эволюция Земли определялась исключительно химическими закономерностями. Около 4 млрд лет назад к ним присоединились биологические закономерности, ставшие последние 0,5 млрд лет ведущими. И, наконец, около двух млн лет назад к двум первым присоединились социальные закономерности, ставшие определяющими в последние 10 тысяч лет. Таким образом, в эволюции Геи выделяются три ствола информации – химический, генетический и культурный и соответственно три сферотемпа.
О семистах миллионах лет истории первого этапа у нас очень мало данных. Главной его чертой является взаимодействие двух протопланет – протоЗемли с протоЛуной. Согласно геофизической модели О. Г. Сорохтина 28 , в начале своей истории Земля представляла холодную, лишенную атмосферы, быстро вращавшуюся (сутки до 3–6 часов) протопланету. Вокруг нее на расстоянии 10 диаметров вращалась расплавленная гравитационным взаимодействием протоЛуна, диск которой, испускавший инфракрасные лучи, в 300–350 раз превышал современный. С периодичностью 18–20 суток протоЗемлю сотрясали землетрясения, энергия которых в 17000 раз превышала современную сейсмоактивность 29 (что и позволяет именовать этот этап сейсмосферотемпом).
Затем, на так называемом пределе Роша 30 , расплавленный металлический материал горба протоЛуны струями был притянут протоЗемлей, а легкая силикатная часть протоЛуны образовала новую планету Луна, которая стала удаляться от Земли. Этот «захват», длившийся не более 10–15 тысяч лет, является первым глобальным кризисом – катастрофой, результатом которой стала «накачка» протоЗемли тепловой энергией (1,25–3,2 10 37 эрг), которая пошла на разогрев Земли. «Захват» кладет начало геологической истории Земли, отделив этап горячей Земли (пирогайю) от этапа холодной Земли (гаплогайи). По мнению О. Г. Сорохтина, приливно-гравитационное взаимодействие двух протопланет ускорило эволюцию Земли (против Венеры) на 2–3 млрд лет (!). В рамках концепции Геи событие на пределе Роша можно трактовать как своеобразное «зачатие» суперорганизма Геи, т. е. формирование условий, необходимых для возникновения жизни. К концу сейсмосферотемпа, к 4,2–4,0 млрд лет, Земля уже имела ядро, обогащенное протолунным железом, и расплавленную мантию, а следовательно, и сильное геомагнитное поле. К этому времени уже полным ходом заработал механизм дегазации, создавший углеродно-мета-ново-паровую атмосферу, плотность которой на порядок превышала современную.
4. Рождение жизни – киральная революция
Киральная чистота – устойчивая дисимметрия оптических стереоизомеров – левоориентированных аминокислот и правоориентированных Сахаров – фундаментальное свойство всего живого. Все мертвые и все космогенные аминокислоты и сахара (в метеоритах) состоят из зеркально симметричных стереоизомеров. Поэтому возникновение киральности и полимеризация как абиогенный синтез – это пограничный рубеж между живым и косным. Причины киральной революции неясны. Одни связывают ее с нарушением четности в слабых взаимодействиях, другие считают результатом разового случая в эволюции. Третьи полагают, что переход от мономеров к полимерам мог происходить только на твердых частицах, т. е. на реголите.
Для возникновения киральности необходимы, очевидно, два условия – наличие свободной воды и сильного геомагнитного поля, т. е. местного, дополнительного к космическому излучению, источника энергии. В Солнечной системе киральность, как теперь ясно, появилась одновременно на двух двойных планетах – Земле и Марсе. Произошло это около 4 млрд лет назад в мелких бассейнах – «лужах-морях», размывавших реголит – космическую пыль, содержащую все биофильные элементы (Н, С, N , О, Р, S ) и минералы – катализаторы. По-видимому, для скопления ориентированных стереоизомеров действительно были благоприятны дефекты кристаллической решетки глинистых минералов, как это и полагает А. Кернс-Смит (1975 г.). Глинистые частицы магнитны и ориентируются вдоль магнитного меридиана. К тому же глины были тогда радиоактивны за счет короткоживушего 26 Al . Таким образом, химическая эволюция практически сразу же дивергировала на Земле на два ствола: гидрогенных минералов – мир симметрии и полимеров – мир асимметрии. В первом стволе эволюционный процесс в форме изоморфных минералов был ограничен уровнем ионных связей элементов, во втором пошел по пути усложнения молекулярных связей, имеющих дипольный момент (Щербаков, 1988). Таким образом, жизнь и геомагнитное поле неотделимы друг от друга.
Появление протожизни – киральности – можно датировать в 3,8–4,0 млрд лет тому назад. А дальше начал действовать механизм обратной связи и отбора на уровне нуклеиново-белковых молекулярных гиперциклов. Как полагает Эйген ( Eigen , 1971), важным фактором самоорганизации протожизни был кооперативный эффект и существование «коллективных» каталических циклов, объединяющих нуклеиновые кислоты и белки. В конкуренции гиперциклов , по-видимому, и появились зачатки генетического аппарата – цепи, построенные из четырех нуклеотидов, сначала РНК (рибонуклеиновой кислоты), а потом и ДНК (дезоксирибонуклеиновой кислоты). Эйген считает, что переход от одиночных макромолекул к каталическому гиперциклу, а от него к закреплению информации в генетическом коде был не менее сложным и многоактовым процессом, чем эволюция от клетки к человеку. Эти процессы, ход которых точно восстановить, конечно, невозможно, совершались уже в первую половину антракогайи, где-то между 4,0–3,6 млрд лет (см. рис. 1).
По мнению М. Нуссинова и В. Харон (1980), В. М. Буреня 31 и С. Л. Шварцева (1997), в механизме возникновения жизни большую роль играли жидкие кристаллы и липидоподобные молекулы жиров. Одновременно с полимеризацией, по-видимому, шло (у пробионтов) и образование мембран – второго фундаментального признака жизни. Следы первых примитивных организмов – пробионтов – в форме миниатюрных трубочек обнаружены сейчас в слоях возрастом в 3,85–3,6 млрд лет. На Земле и на Марсе они разнятся лишь размером. Марсианские пробионты, еще безмембранные, в 100–1000 раз мельче земных 32 .
Третьей фундаментальной особенностью жизни является удивительно тесная связь набора металлов, на которых строятся биохимические процессы земных организмов, с тонкой структурой фотоспектра Солнца. Так, магний, обеспечивающий процессы фотосинтеза и поддерживающий спирали ДНК и РНК, железо, связывающее кислород в крови, натрий и калий, поддерживающие разность электрических потенциалов на стенках клетки, относятся к пяти металлам, которые вместе с кремнием составляют 95% массы солнечной атмосферы. Иными словами, жизнь на Земле зародилась на основе тех металлов, которые характерны для «провалов» фраунгоферовых линий Солнца.
Как отмечают К. Я. Кондратьев и П. П. Федченко, «необходимость обеспечения строго заданной устойчивой геометрии структур молекул – ферментов – одна из причин совпадения металлов, входящих в живые организмы и фотосферу Солнца» 33 .
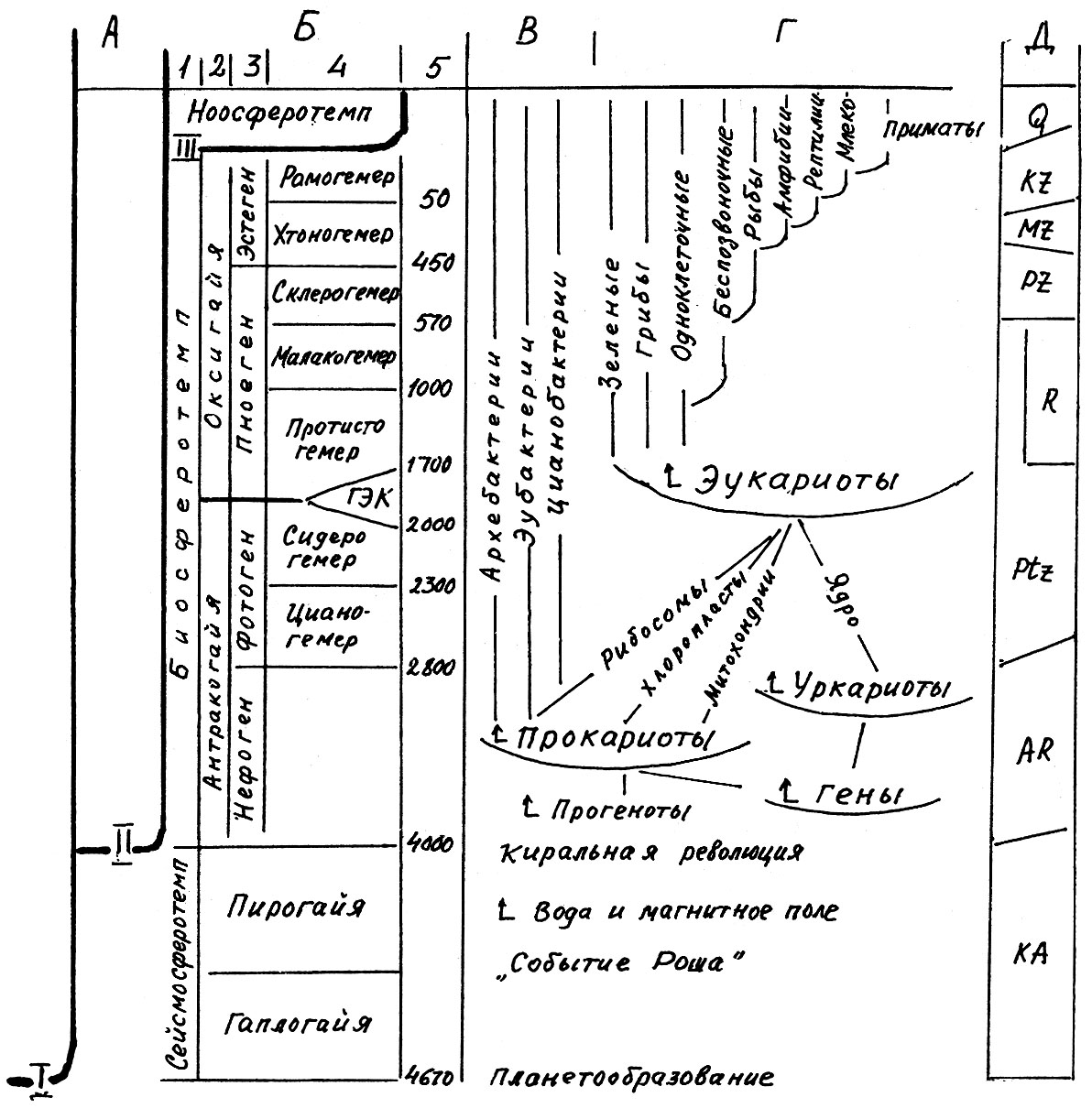
Рис. 1. Темпоральная периодизация эволюции Земли как суперорганизма («Геи»):
А – три информационных ствола эволюции: I – геохимической, II – биогенетической. III социокультурной, каждый из которых начинается с внешнего причинного толчка: плане-тообразования, киральной революции и сексуальной; Б – таксоны темпоральной периодизации («темпы»): 1 - сферотемпы. 2 – тайятемпы, 3 – генотемпы, 4 – гемеротемпы, 5 – начало их в млн лет; В - эволюция прокариотной (доядерной) жизни; Г – схема эндосимбиотического (по Л. Маргулис) становления эукариотной (ядерной) жизни; Д – индексы традиционной хроностратиграфической шкалы (для сравнения)
Сказанное выше позволяет сделать следующие выводы о зарождении жизни: 1) никакого искусственного «засева» жизни на Земле принципиально не могло быть, так как такому «засеву» должно было бы предшествовать и искусственное «создание» Солнца с точно запрограммированными для развития белково-нуклеиновой жизни свойствами фотосферы; 2) возникновение белково-нуклеиновой жизни представляет собой космическое явление, в котором Солнце, Земля и Луна тесно взаимодействуют друг с другом: 3) более того, белково-нуклеиновая жизнь, очевидно, могла возникнуть только на окраинах Галактики, вдали от квазаров с их губительным для жизни жестким излучением.
5. Восстановительная биосфера – «детство Геи» и предрифейская экокатастрофа
В рамках темпоральной периодизации биосферотемп разделя ется на два этапа второго ранга – два гайятемпа (рис. 1). На первом – антракогейском (гр. – углекислый), длившемся 2,3 млрд лет, природная среда имела восстановительный состав. Атмосфера состояла из углекислого газа, азота, метана, аммиака и благородных газов 34 . Кислорода в ней не было, о чем свидетельствует наличие в отложениях того возраста таких кислородофобных минералов, как пирит и уранинит. Химическая эволюция могла происходить только в бескислородной атмосфере, и, следовательно, жизнь могла возникнуть только в восстановительной среде. Антракогайя (ранее автор именовал ее хемогайей) в свою очередь разделяется на два этапа третьего ранга – генотемпа.
Первая половина антракогайи именуется нефогеном (от греч. нефо – облака, ген – родить, т. е. время рожденных под облаками). Земля тогда была окутана непроницаемой для солнечного света мошной паровой атмосферой восстановительного состава, а моря, наоборот, были мелководны. Нефоген был временем существования субклеточных протоорганизмов – прогенот – пробионтов. Они питались за счет первичных запасов восстановительной среды. Вода на поверхности Земли была в то время буквально «живой», так как моря перемывали космогенную пыль – реголит, содержащую все биофильные элементы и минералы – катализаторы. По мере исчерпания запасов реголита появились и более совершенные клеточные доядерные структуры жизни – прокариоты. Мир прокариот, ( Prokaryota ) состоит из двух царств (рис. 1) – архебактерий (метаногенных, галогенных и серозависимых) и эубактерий. Описание их структуры дается в красочно иллюстрированной книге Риса и Стернберга 35 , а более подробно – в монографии Шлегеля (1987). Все прокариоты – это мелкие гаплоидные, т. е. без смены поколений, практически бессмертные организмы, способные обитать в самой разнообразной среде, при высокой температуре и солености. Они представляют собой один из стволов дивергенции жизни, развивающийся по линии отрицательной обратной связи и раз навсегда определенного гомеостаза.
Одновременно возник и другой ствол, развивающийся по линии положительной обратной связи, в которой решающее значение принадлежало принципу экономии энтропии. В этом стволе отбор осуществлялся много быстрее, но возникающие и быстро эволюционирующие организмы не сохранились в ископаемом состоянии. Они получили условное название уркариот (греч. – делающиеся ядерные). Важным элементом организации этого ствола жизни было протоядро, вмешавшее генетические нуклеотидные цепи. Наоборот, в прокариотном стволе главной структурой явились мембраны – инструмент гомеостаза. Поэтому ископаемые остатки прокариот в виде клеточных микросфер довольно часты в отложениях нефогена с возрастом от 3,8 до 3,0 млрд лет (см. рис. 1).
Во вторую половину антракогайи физические условия Земли существенно изменились в связи с возникновением первичного океана. В нем начался грандиозный по масштабам процесс серпентизации – связывания углекислого газа в доломитах и биогенных карбонатных осадках, который предопределил, по А. Б. Ронову (1980), всю последующую химическую эволюцию Земли. Поэтому этот этап (от 2,8 до 1,7 млрд лет) именуется фотогеном (греч. – время рожденных под светом). Так как энергия Солнца была тогда меньше современной на 1/5, то средняя глобальная температура на Земле счала быстро падать от 120–70 °С в конце нефогена до 10–4 °С в конце фотогена (сейчас она 15 °С). Это привело к появлению наземного оледенения – гуронского или гоуганда-грикватаун, датируемого в 2,5–2,2 млрд лет назад (Джон и др., 1982).
Третья бифуркация в истории Земли, в отличие от второй, готовилась долго в условиях так называемого предрифейского экологического кризиса. Датировка его пока не очень точна, по одним данным это 1,7–1,6 млрд лет (Соколов, 1976), по другим 2,2–2,0 млрд лет (Холленд, 1989; Мележик, 1988). Причиной кризиса явилась деятельность бактерий Суапеа – сине-зеленых водорослей. В фотогене они стали лидерами эволюции и были очень широко распространены, образовывали на мелководьях протяженные наслаивающиеся строматолитовые маты (рифы). В качестве продукта своей жизнедеятельности Суапеа, потреблявшие азот, выделяли кислород, бывший ядом для большинства прокариот. Кислород начал накапливаться в замкнутых морях, по Мележику (1988), с 2,2 млрд лет, постепенно губя в них другие прокариоты.
Антракосфера в буквальном смысле слова боролась с этим отравлением путем окисления двухвалентного железа, непрерывно поставлявшегося в океан магматическими излияниями. Вместе с кислородом оно переходило в трехвалентное железо, почти не растворяющееся в воде и поэтому выпадавшее в осадок на дно вместе с карбонатами в форме кремнисто-железистых сезонноленточных (океан был холодным) илов. После метаморфизма из этих илов возникла очень характерная для фотогена порода – джеспилиты, служащие источником сидеритовых руд типа Курской магнитной аномалии. «Солдатами», мобилизованными на борьбу с отравлением кислородом, стали железобактерии, основу которых составляли микроскопические кристаллы сернистого железа. Кстати, за время существования этих бактерий (сидерогемер) на Земле сформировалось 90% всех рудных месторождений. Однако около 2,0 или 1,7 млрд лет назад запасы двухвалентного железа в недрах Земли истощились и кислород стал свободно накапливаться в морях, а потом и в воздухе (Рябенко и др., 1988). Этот момент, именуемый точкой Кюри (концентрация кислорода достигла 10 –3 от современной), хорошо фиксируется появлением на поверхности суши красноцветных ожелезненных песчаников, датируемых в 1,7 млрд лет (Войткевич, Бессонов, 1986; Соколов, 1976).
Некоторые философские выводы:
Первый. Стратегия эволюции восстановительной биосферы, бесспорно, менялась. Ясно выделяются два ее типа – с господством гетеротрофных процессов и автотрофных. На протяжении пефогена органический мир существовал за счет первичных ресурсов – антракосферы. Эта стратегия похожа на вскармливание младенца молоком матери. Фотоген же явно прошел при доминировании аэробных автотрофных процессов питания. При этом ведущим процессом становится потребление азота, а питающиеся им цианобактерии стали лидерами эволюции. Это понятие – лидер эволюции – очень важно для философского осмысления закономерностей эволюции потому, что именно взаимоотношения лидера (неважно, что неразумного) с окружающей средой объективно определяют траекторию и направленность эволюции, в частности соотношение элементов устойчивости и неустойчивости в развитии, движение от гомеостазного равновесия к экологическому кризису.
Второй вывод. На протяжении антракогайи биосфера дважды выходила на грань глобального экологического кризиса и катастрофы. Первый раз это случилось около 2,8 млрд лет назад, когда тектонические процессы вулканизма на Земле достигли максимального за всю ее историю проявления. Резко возрос парниковый эффект атмосферы. А парциальное давление углекислого газа поднялось до максимальной величины в 4,3–4,57 атмосферы 36 . Температура морских вод поднялась до 60–80 °С, опасно приблизясь к закипанию (140 °С). Если бы это случилось, то ситуация на Земле стала бы развиваться по венерианскому типу. Спасло то, что объем воды на Земле быстро увеличивался благодаря дегазации мантии и плитной тектонике. Угроза катастрофы венерианского типа миновала после большой трансгрессии 2,7 млрд лет назад, когда срединно-атлантические хребты, поставлявшие на поверхность Земли основную массу базальтов, оказались под уровнем Океана. Начался спасительный для биосферы процесс серпентизации океанической коры, сопровождавшийся связыванием огромных количеств углекислого газа в доломитах. Благодаря ему к началу фотогена, 2,6 млрд лет назад, давление СО 2 снизилось до 0,7 бар.
Второй раз на грань глобальной экокатастрофы биосфера вышла около 2,2 млрд лет назад, когда единственный тогда на Земле материк Моногея на значительной (а возможно и большей) своей площади покрылся ледниковым щитом. Земля оказалась на грани перехода в «белую планету», т. е. в постоянно замерзшую, наподобие Марса. На этот раз ее спасло начавшееся усиление светимости Солнца (кроме доказанного направленного усиления светимости, оцениваемого в 25–30% за 4 млрд лет, предполагаются и долгопериодические ритмы колебания солнечной активности) 37 .
Третий вывод. Только прерифейский глобальный кризис, вызванный не физическими (в отличие от первых двух), а чисто экологическими причинами, перешел в тотальную экокатастрофу для существовавшей тогда биоты (архебактерий, эубактерий и уркариот). Это подчеркивание философски очень важно. Действительно, накопление в атмосфере только одной тысячной доли от современного содержания кислорода вызвало массовую гибель всех прокариот и уркариот на поверхности материков и океана. Свидетельством гибели являются огромные запасы нефти и газа в рифейских отложениях, образовавшиеся из останков погибшей фауны, и колоссальные по объему толщи графитов рифея. Уцелели лишь те бактерии, которые обитали в сохранившихся азилях восстановительной среды, т. е. в донных отложениях морей, и, конечно, сами цианобактерии.
Таким образом, большая часть экологических ниш на Земле в результате прерифейской тотальной экокатастрофы (ТЭК) опустела, как свидетельствует геологическая летопись, на многие миллионы лет. Вновь жизнь появилась в этих экологических нишах уже в форме высокоорганизованных ядерных организмов ( Eukaryota ), т. е. принципиально нового ствола жизни. Первые их находки датируются в 1,6 и 1,3 млрд лет. Значит ли это, что экологические ниши пустовали 100 или 400 миллионов лет? Не обязательно. Ведущий микропалеонтолог мира Г. Шлегель допускает, что «промежуточные формы не сохранялись и, вероятно, были такими нестойкими, что сейчас нет даже ископаемых остатков, которые бы позволяли судить об их функциональных особенностях... Можно предположить, что на ранних этапах эволюции эукариотической клетки возникали различные модели ее организации, прежде чем появились многоклеточные организмы» 38 .
Появление эукариот (греч. – настоящих ядерных) – величайший скачок в истории жизни. Почти все они аэробы, могут дышать кислородом, а следовательно, получать больше энергии по сравнению с прокариотами, энергетика которых основана на брожении. Клетка эукариот на порядок больше и включает особое ядро, в котором может храниться большой объем генетической информации. Жизненный цикл эукариот включает половой процесс, благодаря которому они способны к быстрой эволюции. Ход ее в общих чертах показан на рис. 1. Таким образом, прерифейская тональная экологическая катастрофа для наземных архебактерий, адаптированных к восстановительной среде, была необходимым условием перехода эволюции на новую, качественно более высокую ступень развития.
Четвертый вывод – о механизме возникновения эукариотического ствола жизни. Хотя детали его, очевидно, навсегда останутся скрытыми от науки, сам принцип его более или менее уже ясен. Это принцип эндосимбиоза, т. е. кооперации уцелевших от гибели мутантов прокариот. Он сформулирован в работах Линн Маргулис 39 и в очень схематичном виде изображен на рис. 1. Исходной нитью к пониманию этого механизма является способность современных эукариот, например амебы, захватывать твердые частицы и живые клетки. Симбиоз, кооперация современных эукариот с прокариотами является образом жизни ядерных организмов. Так, 90% всех растений существует в симбиозе с грибами (на корнях), жвачные животные в симбиозе с бактериями, способными производить ферменты для переваривания целлюлозы, и т. п. Вообще современная биосфера может существовать только в кооперации эукариот и прокариот, автотрофов – фотосинтезаторов кислорода и гетеротрофов – редуцентов органики.
Ясно, что механизм кооперации не складывается поэтапно на протяжении десятков и сотен миллионов лет. Начальным шагом стал эндосимбиоз – «партнерство по выживанию» уркариот с бактериями–мутантами, уже умеющими дышать кислородом (от них хлоропласты у растений и митохондрии у животных) и синтезировать белки (от них рибосомы). Этапы этого партнерства мутантов оказались воплощенными в структуре эукариотической клетки, появившейся в готовом виде где-то около 1,6 млрд лет назад.
6. Земля с кислородной биосферой (оксигайя)
История Земли за последние 1,7 млрд лет и ее традиционные геоисторические подразделения в общих чертах известны всем читателям по школьному курсу биологии. Она предстает в виде «стремительных всплесков эволюции, сменявшихся загадочными массовыми вымираниями, когда за сравнительно короткое время с лица нашей планеты исчезали многие виды живых существ. Однако всякий раз их место занимали новые формы жизни, и некоторые из них оказывались сложнее, чем их предшественники» 40 . Предполагается, что в результате возрастающего обмена информацией, происходящего в процессе полового размножения эукариот, за 1,7 млрд лет в истории оксибиосферы сменилось до 500 млн видов организмов. Однако этот поток информации ученые сейчас склонны считать экзогенным, идущим через вирусы и бактерии 41 . Потому-то филогенетические изменения («мода на мутации») и происходят у разных групп организмов одновременно (фенозоны В. А. Красилова). Поэтому если бы прокариоты сейчас исчезли, то эукариоты не смогли бы существовать самостоятельно, в то время как прокариоты, оставшись одни, могли бы вполне поддерживать существование биосферы, но, разумеется, на примитивном – «восстановительном», антракогейском, уровне.
В рамках темпоральной периодизации оксибиосфера разделяется на «время рожденных дышать» (греч. пноеген) и «время рожденных чувствовать» (эстеген). Дыхание – это решающая адаптация ядерных к отравленной кислородом среде. Процесс дыхания, как известно, высвобождает большее количество энергии – в 35 раз по сравнению с процессом ферментации, используемым прокариотами. И, следовательно, для умеющих дышать открываются гораздо большие возможности прогресса. Но на освоение процесса дыхания у жизни ушло 1,3 млрд лет. Все рубежи внутри оксигеи, или оксигайи (я считаю допустимыми обе транскрипции), определяются скачкообразными изменениями в содержании кислорода в атмосфере.
Так, протистогемер (греч. – день простейших) – время существования только одноклеточных ядерных организмов – характеризуется еще низким, не больше 0,1% от современного, содержанием кислорода. По мнению В. П. Буреня, бифуркация эукариот на одно- и многоклеточных произошла благодаря имеющейся у клеток информации к взаимодействию в популяциях. Эта информация, подобная имеющейся у кристаллов, предопределила прилипание – удержание («адгезирование») части клеток друг к другу 42 . Автор подобный процесс специализации клеток в популяциях назвал цитоколонизацией 43 . Первые следы многоклеточных организмов, в основном червей, появляются в слоях с возрастом 1,1–1,4 млрд лет. Этот рубеж большинством авторов и принимается за точку Кюри – 0,1 % кислорода от современного. Конец «дня мягкотелых» (малакогемера) характеризуется глобально фиксируемым появлением обильной фауны вендского-эдиакарского типа, состоящей из крупных медузовидных организмов, полипов и других кишечнополостных, а также первых бурых водорослей. Вся эта фауна и флора обитала на мелководьях древних морей под защитой десятиметрового слоя воды, спасавшего живое от губительного ультрафиолета.
«День скелетных животных» (гр. склерогемер) начался внезапно 570 млн лет назад появлением первых защищенных панцирем- раковиной, скелетом из карбоната кальция или фосфата организмов – археоциат, брахиопод и трилобитов. Последние были первыми хищниками и первыми животными, у которых есть глаза. Этот кембрийский эволюционный взрыв, когда появилась масса самых различных форм и типов жизни, никак нельзя считать случайным, так же как и появление эдиакарской фауны мягкотелых. Оба «взрыва жизни» произошли вслед за двумя крупными оледенениями, которые нарушили сероводородный режим тогдашних морей и соответствовали скачкам в росте концентрации кислорода в океане. Как этот скачок выражался количественно, неясно. Условно он принимается за точку Пастера – 1–2% от современной его концентрации.
Крупнейшим рубежом в эволюции кислородной биосферы был выход жизни на сушу. Он произошел в конце ордовика – начале силура, между 470 и 420 млн лет назад, опять же вслед за ордовикским оледенением. Толчком к экспансии жизни на сушу стало появление озонового экрана (10% кислорода от современного), обеспечившего защиту от коротковолновой радиации непосредственно на поверхности Земли. Тогда на прибрежных болотах поселились водоросли и плауны, создавшие первые почвы. За ними в реках появились рыбы, которые в новой природной среде приобрели костный позвоночник и челюсти. С них-то и начинается процесс цефализации (от греч. кефале – голова) – роста головы и центральной нервной системы, – ставшей основным направлением прогресса хордовых животных. В темпоральной периодизации весь интервал геологической истории, на котором действует этот процесс, от силура до появления человека, именуется эстегеном (от гр. – чувствовать).
В нем выделяются два поистине революционных события. Первым было появление легких 44 . Процесс его становления начался у двоякодышащих рыб силура и шел 50 млн лет у амфибий (земноводных). Они могли жить и на суше, для этого у них появились лапы и голова, отделенная от плеч, и в воде, где они продолжали откладывать икру. Вторым стало становление класса рептилий (пресмыкающихся), которое началось с «изобретения» амнио-тического (от греч. амнион – мешок) яйца 45 . Хорошо нам знакомое, оно представляет своего рода палеоаналог современного космического аппарата. Эмбрион в нем помещен в мешочек с водной средой, во втором мешочке – запас пищи, желток, в третьем скапливаются отходы эмбриона и, наконец, скорлупа выполняет роль легкого. Таким образом, внутреннее оплодотворение оказалось более надежным способом размножения по сравнению с икрометанием рыб и амфибий. Естественно, что рептилии и птицы в мезозое стали новыми лидерами эволюции.
В отличие от традиционной геоисторической шкалы с семью периодами (рис. 1), я разделяю эстеген всего на два гемеротемпа: хтоногемер (греч. – день Земли) – время экспансии жизни во все природные среды на земле и в воздухе и рамогемер – освоение жизнью лесного биогеоценоза. Я делаю это затем, чтобы отсечь избыточную информацию чисто геоисторического содержания, которая мешает сосредоточиться на прослеживании главной тенденции эволюции биосферы как суперорганизма. Суть ее в переходе от эволюции мозга к эволюции сознания, от цефализации живого к его социализации.
Но прежде отметим два великих вымирания хтоногемера. Первое случилось в конце перми, 250 млн лет назад. Тогда погибло до 50% всех организмов и до 90% обитателей мелководных морей. Причины этого самого грандиозного в оксигайе вымирания не совсем ясны. По мнению автора, оно произошло в результате соединения Гондваны и Лавразии в единый материк – Пангею, резкого сокращения площади шельфовой зоны по этой причине и опреснения океана в результате таяния пермских ледниковых щитов.
Второе вымирание было концом эры динозавров. Оно произошло 65 млн лет назад в результате, как считают сейчас, столкновения Земли с гигантским астероидом, кратер которого, диаметром в 180 км, обнаружен на Юкатане. Последствия столкновения были похожи на «ядерную зиму» – огромные массы пыли минимум на полгода закрыли Солнце и вызвали резкое похолодание, губительное для тропического типа фауны и флоры того времени. Еще страшнее были длительные «кислотные дожди». В этом кризисе, однако, смогли уцелеть некоторые водные рептилии – крокодилы и черепахи – и значительная часть флоры.
Оба великих вымирания нередко причисляются к глобальным экологическим кризисам, но в действительности таковыми не являются. Они не сопровождались какими-либо глобальными геохимическими изменениями, не происходило даже глобальной смены природной зональности.
Экологической революцией хтоногемера явилось появление плаценты у класса млекопитающих – органа, совершенствующего процесс размножения. Поэтому после вымирания высокоспециализированной фауны динозавров 65 млн лет назад место лидеров эволюции заняли млекопитающие.
Однако главный скачок в переходе от эволюции мозга к эволюции сознания произошел позже, в эоцене, между 60–40 млн лет назад, когда на Земле появились приматы – человекообразные обезьяны, адаптированные к самой сложной из всех существующих экологических ниш – дождевым тропическим лесам. Как и все обезьяны, они обладали стереоскопическим зрением, необходимым для прыжков с ветки на ветку, но взамен утраченного в эволюции хвоста получили сильные руки и ладонь с отдельно стоящим большим пальцем. Мозг приматов по сравнению со всеми животными был максимально развит, в особенности в его тыльной и лобных частях. Это, по мнению антропологов 46 , свидетельствует о том, что в жизни высших приматов большую роль играла предтрудовая деятельность. Бесспорно, что использование палок и камней в какой-то мере заменяло им отсутствие клыков и когтей. Бесспорно и то, что высшие приматы обладали достаточно развитой системой биосоциальной информации, передаваемой уже не генетически, а путем подражания, импритинга.
Таким образом, процесс цефализации, начавшийся у речных рыб 450 млн лет назад, доходит у высших приматов до стадии, на которой начинают формироваться контуры принципиально новой, уже надгенетической системы информации. Итог этой направленной эволюции четко отражен в онтогенезе человека – развитии зародыша, в ускоренном виде повторяющем филогенез своих предков.
Говорят, человек произошел от обезьяны. Это далеко не так. Гоминиды отделились от обезьян 30 млн лет назад. Наш же позвоночник начал формироваться еще на стадии рыб. О родстве с ними свидетельствуют и жаберные щели, которые в двухмесячном возрасте трансформируются в горло. Легкие, голову и пятипалую ладонь мы получили в наследство от амфибий и рептилий. Теплую кровь и живорождение – от млекопитающих. От обезьян – хватательную способность рук и стереоскопическое зрение. От высших приматов у нас рудиментарный копчик. И самое главное, наш мозг имеет три слоя: нижний, рудиментарный – наследие стадии пресмыкающихся, средний, центр эмоций, - наследие древних млекопитающих и только верхний слой - кора головного мозга, центр разума формировался на стадии Homo 47 . Таким образом, человек - не творение случая, не творение инопланетного Разума или Бога «по образу своему и подобию», а результат длительной эволюции всей биосферы.
Работа выполнена при финансовой поддержке Фонда Сороса (грант МТ-9000) и Соединенного Фонда Сороса и РФФИ (грант МТ-9300).
(Продолжение следует)
1 Продолжение. Начало см.: Философия и общество. 1998. № 1.
2 Дедю И. И. Экологический энциклопедический словарь. Кишинев. 1990. С. 351.
3 Биология в познании человека/Под ред. Р. С. Карпинской. М., 1989. С. 18.
4 Росляков О. А. Гистерезис платности веществ и его философский смысл. СПб., 1996.
5 Пригожин И., Стенгерс И. Время, хаос, квант. М., 1994.
6 Моисеев Н. Н. Человек и ноосфера. М., 1990.
7 Реймерс Н. Ф. Надежды на выживание человечества. М., 1992.
8 Барановский С. И., Богачев В. Ф., Шевчук Ю. С. Управление будущим. СПб., 1997.
9 Горшков В. Г. Физические и биологические основы устойчивости жизни/ВИНИТИ. М., 1995.
10 Арский Ю. М., Данилов-Данильян В. М., Залиханов М. Ч., Кондратьев К. Я., Котляков В. М., Лосев К. С. Экологические проблемы/МНЭПУ. М., 1997. С. 122, 124.
11 Вернадский В. И. Записка об изучении живого вещества с геохимической точки зрения//Изв. РАН, сер. 6, 1921. Т. 15. С. 120–123; и Научная мысль как планетное явление. М., 1991.
12 Гегамян Г. В. Концепция поля «живого вещества»//Вестн. РАН 1993. Т. 63. № 2. С. 126–133.
13 Гленн Дж. Роль науки и технологии в исследовании будущего//Идеи Н. Д. Кондратьева и динамика общества. М., 1995. С. 217–240.
14 Гольданский В. И. и др. Нарушение зеркальной симметрии и возникновение жизни//Наука и человечество. М., 1986. С. 139–151.
15 Опарин А. И. Жизнь, се природа, происхождение и развитие. М., 1960.
16 Бернал Дж. Возникновение жизни. М., 1969.
17 См.: Хоровиц Н. Поиски жизни в солнечной системе. М., 1988. С. 47.
18 Двойрин Г. В. Единая голографическая информационная теория Вселенной. Научная религия. СПб., 1997.
19 Lovelock J. The ages of Gaia. Biography of our living birth. N. Y. – L. 1988.
20 Камшилов М. М. Эволюция биосферы. М.: Наука, 1974.
21 Горшков В. Г. Физические и биологические основы устойчивости жизни/ВИНИТИ. М., 1995.
22 Шухов В. А., Хон Г. Н. Оправдание случайности. М.: Наука. 1990. 156 с.
23 Пригожий И., Стенгерс И. Время, хаос, квант. М., 1994. Гл. 4.
24 Юзвишин И. И. Информациология. М., 1996. С. 16.
25 Шихобалов Л. С. и Вакуленко А. А. и др. Две статьи в Вестнике СПб., РАЕН. 1997. № 1. С. 369–383.
26 Козырев Н. А. Избр. тр. ЛГУ, 1981.
27 Пригожин И., Стенгерс И. Время, хаос, квант. М., 1994.
28 Сорохтин О. Г. Происхождение Луны и начальные этапы эволюции Земли//Жизнь на Земле. М., 1988. С. 5–23.
29 Сорохтин О. Г., Ушаков С. А. Глобальная эволюция Земли. М.: Изд-во МГУ, 1991. 446 с.
30 Расстояние в десять диаметров Земли, на котором сила притяжения протоЗемли равна силе тяжести на протоЛуне.
31 Природа и дух. Мир философских проблем. СПб., 1995. С. 23–26.
32 Сэвидж Д., Хартсфилд Д., Солсбери Д. Жизнь на Марсе?//3емля и Вселенная. 1997. № 1. С. 50.
33 Кондратьев К. Я., Федченко П. П. Тонкая структура спектра Солнца и ее роль в эволюции биосферы. СПб., 1992. С. 16.
34 По данным Л. И. Салона (Общая стратиграфическая шкала докембрия. М.: Недра. 1973), воздушные пузырьки в катархейских кварцитах представлены на 60% углекислым газом и на 35% разными «кислыми дымами», в долях пропета азотом и благородными газами, кислорода нет вовсе.
35 Рис Э., Стернберг Э. От клеток к атомам. М.: Мир. 1988. 143 с.
36 Сорохтин О. Г., Ушаков С. А. Происхождение и эволюция атмосферы Земли//Жизнь Земли. 1992. С. 5–44.
37 В океане СО2 сейчас содержится в 57 раз больше, чем в атмосфере, а в горных породах – в 70 раз (Монин А. С. История Земли. Л.: Недра, 1977. С. 66). Начиная с 2,7 млрд лет в этом направлении и пошло перераспределение химического состава геосфер Земли.
38 Шлегель Г. Общая микробиология. М.: Мир. 1987. С. 524–525.
39 Маргулис Л. Роль симбиоза в эволюции клетки. М., 1983. 351 с.
40 Бейли Д., Седдон Т. Доисторический мир. М.: Росмэн, 1995. С. 6.
41 Маргулис Л. Там же: Красилов В. А. Эволюция и биостратиграфия. М.: Наука. 1977: Горшков В. Г. Физические и биологические основы устойчивости жизни. М., 1995. С. 182.
42 Природа и дух. СПб., 1995. С. 29.
43 Зубаков В. А. XXI век, сценарии будущего. 1995. С. 12. Аналогией его в какой-то мере могут быть физиологические процессы, и ныне идущие в колониях слизевиков-миксомицет. Большую часть своей жизни они проводят в виде отдельных клеток, питаясь бактериями на коре деревьев. Когда же запас пищи у них иссякает, миллионы их собираются, очевидно, в ответ на химическую информацию, в одну клеточную массу, как бы переходящую в новое качество многоклеточного организма: Он передвигается, реагирует на внешнюю обстановку и. как гриб, выбрасывает плодоножку – спорангий. Таким образом, временное объединение слизевиков в многоклеточный организм есть форма выживания в ответ на кризис питания. После того как споры разлетятся по окрестностям и, стало быть, экспансия будет достигнута, организм умирает (Доисторический мир. М., 1995. С. 54).
44 Легче всего осуществили переход с моря на сушу членистоногие, а именно насекомые, обладающие способностью дышать всей поверхностью тела. Но летать они научились только в карбоне. См.: Флинт Р. Ф. История Земли. М.: Прогресс, 1978.
45 Флинт Р. Ф. Там же. С. 209.
46 Нестурх М. Ф. Происхождение человека. М.: Наука, 1970.
Размещено в разделах




